














Published online May 26, 2026. doi: 10.4252/wjsc.v18.i5.114668
Revised: November 25, 2025
Accepted: March 5, 2026
Published online: May 26, 2026
Processing time: 242 Days and 15.4 Hours
Extracellular vesicles (EVs) are small particles that range between 30-150 nano
To perform a proteomic analysis of EVs from bone marrow-derived mesenchymal stem cells (BMSCs) and to identify core and unique protein signatures of BMSC-derived EVs, explore the enriched pathways and biological processes they influence, and highlight potential biomarkers or therapeutic targets relevant to regenerative medicine and disease modulation.
EVs were isolated from mesenchymal stromal cells derived from the bone marrow of healthy male donors using standard techniques. EVs were identified using sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Protein was isolated from the EVs and subjected to liquid chromatography and mass spectrometry analysis using liquid chromatography and mass spectrometry detection. Metascape (https://metascape.org/) was used to perform Gene Ontology annotation and Kyoto Encyclopedia of Genes and Genomes pathway enrichment analysis of common proteins in all samples. A minimum overlap of ≥ 2 proteins and a P-value < 0.01 was considered statistically significant. The STRING database was utilized to predict protein-protein interactions for common proteins and intersecting proteins involved in diseases.
Proteomic analysis revealed that 180 proteins were recognized in sample one, 130 in sample two, 113 in sample three, and 170 in sample four. Fifty-three proteins were identified, and their sizes ranged from 11 kDa to 245 kDa. Among all four BMSC-derived EV samples, 53 proteins were consistently present across all samples. The unique protein signatures may reflect donor-specific or culture-specific differences and could represent potential biomarkers for patient stratification if validated in larger datasets. Fifteen of the identified proteins were found to be related to regenerative medicine, and five proteins very important in neurological disease/disorder relevance.
Our study reveals that different proteins can be identified and separated from BMSCs, providing a window of opportunity to test the functionality of EVs in trials without directly using BMSCs. It remains to be seen whether these individual EVs will exert the same physiological levels as BMSCs in influencing the behavior of recipient cells in modifying disease processes, affecting the immune system, and facilitating tissue regeneration.
Core Tip: In regenerative medicine, mesenchymal stem cell (MSC)-based therapy is used to repair diseased or damaged tissue. The technology of culturing MSCs is expensive and not available everywhere. MSC-derived extracellular vesicles have gained attention as a potential therapeutic alternative to MSCs themselves. Therefore, scientists should be able to isolate and identify individual proteins and their functionality, which can be useful for therapeutic purposes. We used proteomics to isolate and identify 53 proteins from MSC-derived extracellular vesicles, 15 of which can be targeted in the field of regenerative medicine.
- Citation: Sadat-Ali M, Al-Turki HA, Khan AA, Bannu SM, Vutharadhi S, Baig MM. Isolation and proteomic profiling of mesenchymal stem cell-derived extracellular vesicles: Unlocking new frontiers in regenerative and translational medicine. World J Stem Cells 2026; 18(5): 114668
- URL: https://www.wjgnet.com/1948-0210/full/v18/i5/114668.htm
- DOI: https://dx.doi.org/10.4252/wjsc.v18.i5.114668
Mesenchymal stem cells (MSCs) are a type of multipotent adult stem cell with the capacity to self-renew and differentiate into several lineages, including bone, cartilage, muscle, and adipocytes. The benefits of regenerative medicine are so immense that now many diseases are being treated in many countries. According to a report published by the National Institutes of Health, there are over 8000 stem cell products being tested in clinical trials[1]. Specifically, ClinicalTrials.gov has registered 6205 clinical trials on stem cell therapy worldwide as of May 18, 2021, with 1240 focused on bone marrow-derived MSC (BMSC) therapy[2]. MSCs can repair tissues in regeneration through immunomodulation, differentiating into specialized cells, and secreting growth factors[3-5]. Initial MSC research focused on the multilineage differentiation of cells towards desired phenotypes. It has been recognized that many of the therapeutic effects of MSCs are mediated through the release of exosomes and extracellular vesicles (EVs). EVs represent a broad category of lipid bilayer-enclosed vesicles released by cells, whereas exosomes are a specific subtype of EVs. Proteins secreted by MSCs contribute to the formation of a microenvironment that promotes tissue repair at sites of injury[6]. These MSC-mediated effects are particularly important in the treatment of tissue damage caused by vascular interruption, as they promote vascularization and restore perfusion through angiogenesis, which is essential for preventing fibrosis.
MSCs are known for their regenerative capabilities and immunomodulatory properties and have demonstrated therapeutic potential in tissue repair, inflammation, and immune regulation[3]. MSCs exert their effects through both their cell membrane and intracellular components. The MSC membrane consists of a lipid bilayer containing phos
Exosomes are membrane-bound vesicles that originate from the endosomal network within cells, whereas other EVs, such as microvesicles, are formed through direct budding from the plasma membrane. Although all exosomes are EVs, not all EVs are exosomes[9]. These vesicles, typically ranging from 30-150 nanometers in diameter, carry a diverse cargo of proteins, lipids, and nucleic acids, enabling them to influence the function of recipient cells[10,11].
EVs typically range from approximately 50 nm to 1 μm in diameter and carry a diverse cargo of proteins, lipids, and nucleic acids, enabling them to function as intercellular communication mediators that influence the behavior of recipient cells. EVs derived from MSCs (MSC-EVs) have shown promise in mimicking many of the beneficial effects of MSCs, offering a cell-free therapeutic approach with reduced immunogenicity. These vesicles promote tissue regeneration, modulate immune responses, and have potential applications in the treatment of various diseases, including cancer[12,13].
Despite increasing interest in MSC-EVs, significant gaps remain in our understanding of their proteomic landscape. This knowledge gap highlights both the therapeutic potential of EVs and the possibility of developing chemically synthesized EV-like systems[14,15]. Although several studies have profiled EV cargo, comprehensive analyses of their protein composition, including comparative, functional, and pathway-specific proteomics across different conditions or cellular sources, remain limited[16,17]. In addition, standardized methodologies for EV isolation and characterization are still lacking, which contributes to variability and complexity across studies[18].
The aim of this study was to perform a comprehensive proteomic analysis of EVs derived from BMSCs across five different biological or experimental conditions. Through this approach, we sought to identify both core and condition-specific protein signatures of BMSC-derived EVs, explore the enriched pathways and biological processes they regulate, and highlight potential biomarkers or therapeutic targets relevant to regenerative medicine and disease modulation.
The present study was approved by the Ethics Committee of Stem Cells Regenerative and Research Labs Inc., Towli Chowki, Hyderabad, Telangana, India, (Approval No. 10296/2025). All experiments were conducted in accordance with the guidelines of the Declaration of Helsinki.
Informed consent was obtained from 4 male participants for bone marrow aspiration, and permission was granted to publish the data derived from these investigations. All patients underwent detailed clinical examination and relevant blood investigations to rule out underlying disease conditions. The mean age of the bone marrow donors was 33.25 ± 5.3 years.
Bone marrow aspiration was performed using an 11-gauge Jamshidi needle pre-flushed with heparin (100000 units in 10 mL of normal saline). Approximately 30 mL of bone marrow was aspirated and transferred into a 50 mL tube con
MSCs were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM) supplemented with 10% fetal bovine serum and 1% penicillin-streptomycin solution in a humidified incubator at 37 °C with 5% CO2. The culture medium was replaced every 3-4 days. When the cells reached approximately 80% confluence, they were harvested for protein extraction. To prevent contamination, cultures were periodically treated with Fungizone and BM-Cyclin to inhibit fungal and mycoplasma growth.
Serum-free conditioned media from MSC cultures was collected and transferred to 50 mL conical tubes. Cells were trypsinized and assessed for viability, which was approximately 1.5 × 106 cells with ≥ 95% viability. The conditioned media was centrifuged at 3000 × g for 20 minutes at 4 °C to remove cell debris and apoptotic bodies, followed by filtration through a 0.22 μm pore membrane filter. The resulting cell-free supernatant was transferred to open-top, thin-wall ultracentrifuge tubes and centrifuged at 100000 × g for 1 hour at 4 °C using an SW32 Ti rotor. After ultracentrifugation, the supernatant was carefully removed using a Pasteur pipette. The EV pellet was resuspended in 200 μL of freezing medium and stored at -80 °C for subsequent use within 24 hours.
Exosomal proteins were analyzed using 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE, Invitrogen™, Thermo Fisher Scientific, NY, United States). Protein samples were loaded onto the gel and separated at 120 V for 60 minutes in MOPS SDS running buffer. A prestained protein marker (Bio-Rad, Cat. No. 1610374, CA, United States) was included in one lane to monitor protein migration. Following electrophoresis, the gels were stained with Coomassie Brilliant Blue (Bio-Rad, Cat. No. 1610786, CA, United States) to visualize protein bands. Western blotting was subsequently performed to confirm the presence of exosomal markers (CD9, CD63, CD81) and the absence of the cellular contamination marker calnexin.
Exosomes were lysed using a combination of chemical and physical methods. Vesicles were suspended in a lysis buffer containing 8 M urea to denature proteins and 1% protease inhibitor to prevent protein degradation. The suspension was then sonicated three times at high intensity to disrupt the exosomal membranes. Following lysis, the protein extract was centrifuged at 12000 × g for 15 minutes at 4 °C to remove insoluble debris. The clarified supernatant containing the solubilized proteins was carefully collected. Total protein concentration was measured using a bicinchoninic acid protein assay kit (Thermo Fisher Scientific, Cat. No. 85165, Waltham, MA, United States) according to the manufacturer’s instructions.
Following quantification, protein samples were prepared for mass spectrometry analysis. Proteins were first reduced with 5 mmol/L dithiothreitol for 30 minutes at 56 °C. Free sulfhydryl groups were then alkylated with 11 mmol/L iodoacetamide for 15 minutes at room temperature in the dark. The urea concentration was subsequently diluted to < 2 M using 100 mmol/L triethylammonium bicarbonate buffer to ensure optimal trypsin activity. Proteins were enzymatically digested into peptides using trypsin. A two-step digestion protocol was employed: First, trypsin was added at a 1:50 enzyme-to-protein mass ratio and incubated overnight at 37 °C; this was followed by a second digestion with trypsin at a 1:100 ratio for an additional 4 hours. The resulting peptides were desalted using C18 Ziptips (Millipore, Darmstadt, Germany) to remove salts and other contaminants. Purified peptides were then eluted with 0.1% trifluoroacetic acid in 50%-70% acetonitrile, lyophilized, and reconstituted in 1% formic acid and 5% acetonitrile prior to mass spectrometry analysis.
Digested peptide samples were analyzed by liquid chromatography-mass spectrometry (LC-MS/MS) as previously described by Li et al[19]. Peptides were separated by reverse-phase liquid chromatography using an EASY-nLC 1000 UPLC system and analyzed on a Q Exactive™ Plus mass spectrometer (Thermo Fisher Scientific, Waltham, MA, United States). The chromatographic gradient consisted of a linear increase from 6% to 80% solvent B over 40 minutes at a flow rate of 400 nL/minute. For data-independent acquisition mass spectrometry (DIA-MS), a full MS scan (m/z 350-1500, resolution 120000) was followed by DIA scans using 50 variable isolation windows. DIA raw files were processed using Spectronaut X, with iRT-based retention time prediction and local normalization applied during data analysis. Peptide precursors and proteins were filtered at a false discovery rate (FDR) of 1% using the mProphet algorithm. Protein intensity was calculated based on the summed peak areas of MS2 fragment ions. The FDR was estimated using the ratio of decoy hits (false positives) to the total number of target and decoy identifications according to the following formula:
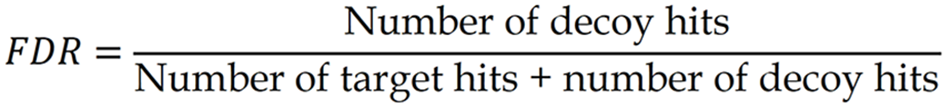 .
.
Further processing was performed using ProteinLynx Global Server software (Waters Corporation). Protein abundance read counts were normalized using the total number of reads mapped to Sequin spike-in controls or RC spike-in controls as size factors. Comparative analyses were conducted across the MSC-EV datasets to identify shared and common proteins.
Metascape (https://metascape.org/) was used to perform Gene Ontology (GO) annotation and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis of common proteins found in all samples. A minimum overlap of ≥ 2 proteins and a P-value < 0.01 was considered statistically significant.
DAVID (version 6.8) and Metascape software were utilized to analyze the pathways enriched among the common proteins. Key pathways were selected to understand disease pathogenesis and identify reliable drug prediction targets. This was followed by pathway network analysis of disease-related genes, which revealed a network among different genes related to the Tree tab.
The STRING database was used to predict protein-protein interactions (PPIs) among common and disease-associated intersecting proteins. The resulting interaction networks were imported into Cytoscape software (version 3.8.2), and hub genes were identified using the cytoHubba plugin with the Maximal Clique Centrality (MCC) algorithm. The top 10 hub genes were ranked in descending order according to their MCC scores, with the highest-scoring genes considered the most significant hub genes.
Figure 1 shows MSCs cultured on days 2 and 19. The average retention time of the maximum chromatographic peak was 32.21 ± 9.09 seconds. Each sample exhibited a minimum of 12 peaks, with each peak corresponding to a distinct compound. Proteins were identified based on the position of the peaks according to their retention times. Figure 2 presents the chromatograms obtained from the four donors. Among these chromatograms, three exhibited similar peak intensities, indicating comparable compound concentrations. SDS-PAGE was performed to separate proteins based on molecular weight and to visually verify the sizes of the identified proteins. This analysis confirmed that the proteins detected by liquid chromatography-mass spectrometry were present in the EV samples. The protein profile obtained from LC-MS/MS analysis was consistent with the expected physical properties of proteins found in EVs. The identified proteins ranged from 11 kDa to 245 kDa (Figure 3).



Based on the established criteria and analysis, a total of 53 proteins were identified at a protein-level FDR of < 1% and a P-value < 0.05. The presence of EVs was further confirmed by Western blotting, which demonstrated the presence of exosomal markers (CD9, CD63, CD81) and the absence of the cellular contamination marker calnexin. EV morphology was also validated using transmission electron microscopy (Figure 4). Subsequent analyses explored the functional roles and interconnections of the identified EV proteins. Protein-protein interaction network analysis and functional enrichment analysis were performed to identify biologically meaningful associations among these proteins (Figure 5).


Proteomic analysis of EVs isolated from MSCs revealed variable numbers of proteins across the four biological samples. A total of 180 proteins were identified in sample 1, 130 in sample 2, 113 in sample 3, and 170 in sample 4. The variation in protein counts across replicates likely reflects donor-specific heterogeneity as well as technical variability in mass spectrometry sensitivity (Figure 6).

To identify conserved proteins, we performed Venn diagram analysis. Across all four MSC-EV samples, 53 proteins were consistently detected. These shared proteins likely represent the core MSC-EV proteome and may play fundamental roles in vesicle biogenesis, cargo trafficking, and intercellular communication. The identification of this conserved protein set increases confidence in their biological relevance compared with proteins uniquely detected in individual samples.
In addition to the 53 conserved proteins (Table 1), each sample contained a subset of unique proteins that were not detected in the other replicates. Sample 1 exhibited the highest number of unique proteins, whereas sample 3 showed the lowest. These unique protein signatures may reflect donor-specific or culture-specific differences and could represent potential biomarkers for patient stratification if validated in larger datasets. Table 2 lists the proteins associated with regenerative medicine and 5 proteins with particular relevance to neurological diseases and disorders. Based on the applied criteria and analysis, a total of 53 were identified at a protein-level FDR of < 1% protein and a P-value < 0.05.
| No. | UniProt ID(s) | Protein/gene | Protein name | Function |
| 1 | Q7Z460 | CLASP1 | CLIP-associating protein 1 | Stabilization and polarization of cytoplasmic microtubules |
| 2 | O75122 | CLASP2 | CLIP-associating protein 2 | Mediator of ERBB2-dependent stabilization of microtubules at the cell cortex |
| 3 | Q86SQ0 | PHLDB2 | Pleckstrin homology-like domain family B member 2 | Plays a role in acetyl-choline receptor aggregation in the postsynaptic membrane |
| 4 | Q32MQ0 | ZNF750 | Zinc finger protein 750 | Transcription factor involved in epidermis differentiation |
| 5 | Q6NXG1 | ESRP1 | Epithelial splicing regulatory protein 1 | mRNA splicing factor that regulates the formation of epithelial cell-specific isoforms, regulates splicing and expression of genes involved in inner ear development, auditory hair cell differentiation, and cell fate specification in the cochlear epithelium |
| 6 | O94880 | PHF14 | PHD finger protein 14 | PDGFRA expression, regulates mesenchymal cell proliferation, suppresses the expression of CDKN1A/p21 by reducing the level of trimethylation of histone H3 ‘Lys-4’, leading to enhanced proliferation of germinal center B cells |
| 7 | P17844 | DDX5 | Probable ATP-dependent RNA helicase DDX5 | Transcriptional coactivator for RUNX2 and involved in regulation of osteoblast differentiation, control of the circadian rhythms |
| 8 | Q92841 | DDX17 | Probable ATP-dependent RNA helicase DDX17 | Involved in multiple cellular processes, including pre-mRNA splicing, alternative splicing, ribosomal RNA processing and miRNA processing, as well as transcription regulation |
| 9 | Q8WWZ7 | ABCA5 | Cholesterol transporter ABCA5 | Plays a role in the processing of autolysosomes |
| 10 | Q6W2J9 | BCOR | BCL-6 corepressor | Acts as a negative regulator of osteo-dentinogenic capacity in adult stem cells |
| 11 | Q5SW24 | DACT2 | Dapper homolog 2 | Regulates intracellular signaling pathways during development, control of morphogenetic behavior of kidney ureteric bud cells |
| 12 | Q13127 | REST | RE1-silencing transcription factor | Plays a role in repression of miR-132 expression in hippocampal neurons, thereby leading to neuronal cell death |
| 13 | Q8N163 | CCAR2 | Cell cycle and apoptosis regulator protein 2 | Maintains genomic stability and cellular integrity following UV-induced genotoxic stress |
| 14 | Q13263 | TRIM28 | Transcription intermediary factor 1-beta | Maintains a transcriptionally repressive state of genes in undifferentiated embryonic stem cells, required for activated KRAS-mediated promoter hypermethylation and transcriptional silencing of tumor suppressor genes or other tumor-related genes in colorectal cancer cells |
| 15 | Q9UPY3 | DICER1 | Endoribonuclease dicer | Plays a central role in short dsRNA-mediated post-transcriptional gene silencing |
| 16 | Q14999 | CUL7 | Cullin-7 | Regulates Golgi morphogenesis and dendrite patterning in brain, acts as a regulator in trophoblast cell epithelial-mesenchymal transition and placental development |
| 17 | Q8TCJ2 | STT3B | Dolichyl-diphosphooligosaccharide-protein glycosyltransferase subunit STT3B | Plays a role in endoplasmic reticulum-associated degradation pathway that mediates ubiquitin-dependent degradation of misfolded endoplasmic reticulum proteins |
| 18 | Q2KJY2 | KIF26B | Kinesin-like protein KIF26B | Helps in embryonic kidney development |
| 19 | Q9NSC2 | SALL1 | Sal-like protein 1 | Transcriptional repressor involved in organogenesis. Plays an essential role in ureteric bud invasion during kidney development |
| 20 | Q7Z591 | AKNA | Microtubule organization protein AKNA | Centrosomal protein that plays a key role in cell delamination by regulating microtubule organization |
| 21 | Q9Y4H2 | IRS2 | Insulin receptor substrate 2 | Plays role in development, growth, glucose homeostasis as well as lipid metabolism |
| 22 | P35568 | IRS1 | Insulin receptor substrate 1 | Plays important role in development, growth, glucose homeostasis as well as lipid metabolism |
| 23 | P42229 | STAT5A | Signal transducer and activator of transcription 5A | Regulates the expression of milk proteins during lactation |
| 24 | P22681 | CBL | E3 ubiquitin-protein ligase CBL | Participates in signal transduction in hematopoietic cells, regulates osteoblast differentiation and apoptosis |
| 25 | P42224 | STAT1 | Signal transducer and activator of transcription 1-alpha/beta | Signal transducer and transcription activator that mediates cellular responses to interferons, cytokine KITLG/SCF and other cytokines and other growth factors |
| 26 | Q8TDM6 | DLG5 | Disks large homolog 5 | Acts as a regulator of the Hippo signaling pathway, regulating cell proliferation, maintenance of epithelial polarity, epithelial-mesenchymal transition, cell migration and invasion, in dendritic spine formation and synaptogenesis in cortical neurons |
| 27 | Q9UK61 | TASOR | Protein TASOR | Multiprotein complex that mediates epigenetic repression, plays a crucial role in early embryonic development, in maintaining epiblast fitness or potency |
| 28 | Q9BRK4 | LZTS2 | Leucine zipper putative tumor suppressor 2 | Required for central spindle formation and the completion of cytokinesis |
| 29 | O60293 | ZFC3H1 | Zinc finger C3H1 domain-containing protein | Subunit of the trimeric poly(A) tail exosome targeting complex that directs a subset of long and polyadenylated poly(A) RNAs for exosomal degradation |
| 30 | Q9H7Z3 | NRDE2 | Nuclear exosome regulator NRDE2 | Responsible for DNA damage |
| 31 | Q13009 | TIAM1 | Rho guanine nucleotide exchange factor TIAM1 | Guanyl-nucleotide exchange factor that activates RHO-like proteins and connects extracellular signals to cytoskeletal activities |
| 32 | Q9H4I2 | ZHX3 | Zinc fingers and homeoboxes protein 3 | Acts as a transcriptional repressor, involved in the early stages of mesenchymal stem cell osteogenic differentiation |
| 33 | Q93052 | LPP | Lipoma-preferred partner | Plays a structural role at sites of cell adhesion in maintaining cell shape and motility |
| 34 | Q7Z7 L1 | SLFN11 | Schlafen family member 11 | Specifically abrogates the production of retroviruses such as human immunodeficiency virus 1 by acting as a specific inhibitor of the synthesis of retroviruses encoded proteins in a codon-usage-dependent manner |
| 35 | P53804 | TTC3 | E3 ubiquitin-protein ligase TTC3 | Regulates neuronal differentiation by regulating actin remodeling and Golgi organization via a signaling cascade involving RHOA, CIT and ROCK |
| 36 | Q16363 | LAMA4 | Laminin subunit alpha-4 | Mediates the attachment, migration and organization of cells into tissues during embryonic development |
| 37 | P26006 | ITGA3 | Integrin alpha-3 | Participates in the adhesion, formation of invadopodia and matrix degradation processes, promoting cell invasion |
| 38 | P02452 | COL1A1 | Collagen alpha-1 (I) chain | Group I collagen (fibrillar forming collagen) |
| 39 | P16234 | PDGFRA | Platelet-derived growth factor receptor alpha | Regulates embryonic development, cell proliferation, survival and chemotaxis, differentiation of bone marrow-derived mesenchymal stem cells |
| 40 | P46934 | NEDD4 | E3 ubiquitin-protein ligase NEDD4 | Part of a signaling complex composed of NEDD4, RAP2A and TNIK which regulates neuronal dendrite extension and arborization during development |
| 41 | P17813 | ENG | Endoglin | Regulates angiogenesis, migration of vascular endothelial cells |
| 42 | Q15678 | PTPN14 | Tyrosine-protein phosphatase non-receptor type 14 | Regulates lymphangiogenesis, cell-cell adhesion, cell-matrix adhesion, cell migration, cell growth and also regulates TGF-beta gene expression |
| 43 | P29320 | EPHA3 | Ephrin type-A receptor 3 | Upon activation by EFNA5, it regulates cell-cell adhesion, cytoskeletal organization and cell migration; in cardiac cells, it controls migration and differentiation and regulates the formation of the atrioventricular canal and septum |
| 44 | P54764 | EPHA4 | Ephrin type-A receptor 4 | Plays an important role in the development of the nervous system by controlling different steps of axonal guidance; in axonal guidance, it plays a role in synaptic plasticity |
| 45 | Q9NQC3 | RTN4 | Reticulon-4 | Induces the formation and stabilization of endoplasmic reticulum tubules, inhibits spreading, migration and sprouting of primary brain microvascular endothelial cells, stabilization of wiring and restriction of plasticity in the adult central nervous system |
| 46 | Q13464 | ROCK1 | Rho-associated protein kinase 1 | Protein kinase that is a key regulator of the actin cytoskeleton and cell polarity, involved in osteoblast compaction, may regulate closure of the eyelids and ventral body wall |
| 47 | O75116 | ROCK2 | Rho-associated protein kinase 2 | Protein kinase that is a key regulator of actin cytoskeleton and cell polarity. Regulates smooth muscle contraction, actin cytoskeleton organization, stress fiber and focal adhesion formation, neurite retraction, cell adhesion and mortality |
| 48 | Q9P2M7 | CGN | Cingulin | Plays a role in the formation and regulation of the tight junction paracellular permeability barrier |
| 49 | Q68DE3 | USF3 | Basic helix-loop-helix domain-containing protein USF3 | Involved in the negative regulation of epithelial-mesenchymal transition |
| 50 | Q9NUX5 | POT1 | Protection of telomeres protein 1 | Component of the telomerase ribonucleoprotein complex that is essential for the replication of chromosome termini |
| 51 | Q6EMK4 | VASN | Vasorin | Expressed at highest levels in aorta, at intermediate levels in kidney and placenta and at lowest levels in brain, heart, liver, lung and skeletal muscle |
| 52 | Q9C0F0 | ASXL3 | Putative polycomb group protein ASXL3 | The PR-DUB complex is an epigenetic regulator of gene expression and acts as a transcriptional coactivator, affecting genes involved in development, cell communication, signaling, cell proliferation and cell viability |
| 53 | Q8IXJ9 | ASXL1 | Polycomb group protein ASXL1 | Together with BAP1, negatively regulates epithelial-mesenchymal transition of trophoblast stem cells during placental development by regulating genes involved in epithelial cell integrity, cell adhesion and cytoskeletal organization. Probable polycomb group protein involved in transcriptional regulation mediated by ligand-bound nuclear hormone receptors, such as retinoic acid receptors and peroxisome proliferator-activated receptor gamma |
| Category | UniProt ID(s) | Gene symbol | Protein name | Regenerative function |
| ECM remodeling and structural support | P02452 | COL1A1 | Collagen type I alpha-1 | Structural ECM, Wound healing, tissue strength |
| P35568 | FLNA | Filamin-A | Cytoskeletal remodeling, motility, tissue repair | |
| Q16363 | ITGA6 | Integrin alpha-6 | Stem cell adhesion, marker of regenerative potential | |
| Q93052 | ITGB1BP1 | Integrin beta-1-binding protein 1 | Integrin signaling in wound healing | |
| Growth factor and regenerative signaling | P16234 | PDGFRB | PDGF receptor beta | Angiogenesis, proliferation, repair |
| P29320 | SHC1 | SHC-transforming protein 1 | Growth factor adapter, survival, regeneration | |
| P37275 | PPARG | Peroxisome proliferator-activated receptor gamma | Differentiation, tissue remodeling | |
| P42224 | STAT1 | STAT1 | JAK/STAT repair and immune regulation | |
| P42229 | STAT2 | STAT2 | Anti-viral and repair signaling | |
| Q13009 | SMAD3 | SMAD3 | TGF-β pathway, fibrosis, repair | |
| Q13127 | SMAD4 | SMAD4 | Central TGF-β/BMP signaling in regeneration | |
| Chaperones and stress response | Q13464 | HSP90AA1 | Heat shock protein HSP90α | Protects stem cells, enhances survival in stress |
| Cell proliferation and stemness regulators | Q14999 | NPM1 | Nucleophosmin | Proliferation, ribosome biogenesis, stem cell support |
| Migration and tissue remodeling | P46934 | IQGAP1 | IQGAP1 | Cell migration, cytoskeletal organization, repair |
| Q92841 | LRP1 | LDL receptor related protein 1 | Neuro regeneration, ECM clearance, repair |
To characterize the functional landscape of MSC-EV proteins, we performed GO and KEGG pathway enrichment analyses using the experimentally identified proteome. Across all GO domains, cellular component, molecular function, and biological process, we observed significant enrichment patterns supported by robust statistical parameters, including fold enrichment, P-values, FDR-adjusted P-values, and the number of contributing proteins (Table 3).
| Term ID | Term description | Observed protein count | False discovery rate |
| hsa04510 | Focal adhesion | 7 | 0.00066 |
| hsa05205 | Proteoglycans in cancer | 7 | 0.00066 |
| hsa04530 | Tight junction | 6 | 0.00091 |
| hsa05206 | MicroRNAs in cancer | 6 | 0.00091 |
| hsa05200 | Pathways in cancer | 8 | 0.0092 |
| hsa04810 | Regulation of actin cytoskeleton | 5 | 0.0229 |
| hsa04935 | Growth hormone synthesis, secretion and action | 4 | 0.0229 |
GO cellular component analysis revealed that MSC-EV proteins were strongly enriched in extracellular exosomes, EVs, and membrane-bounded vesicles (FDR range: 2.3 × 10-4 to 3.9 × 10-3). These terms exhibited high fold-enrichment values (up to approximately 6-fold) and substantial protein counts (ranging from 2 to 57), as shown in Table 4. These findings are consistent with the vesicular origin of the isolated samples (Figure 7A).

| Term ID | Term description | Observed protein count | False discovery rate |
| GO:0005622 | Intracellular anatomical structure | 57 | 0.0027 |
| GO:0043229 | Intracellular organelle | 54 | 0.0027 |
| GO:0045180 | Basal cortex | 3 | 0.0027 |
| GO:0043226 | Organelle | 55 | 0.0033 |
| GO:0043232 | Intracellular non-membrane-bounded organelle | 31 | 0.0045 |
| GO:0005654 | Nucleoplasm | 26 | 0.014 |
| GO:0043227 | Membrane-bounded organelle | 52 | 0.0153 |
| GO:0030981 | Cortical microtubule cytoskeleton | 2 | 0.0261 |
| GO:0032991 | Protein-containing complex | 30 | 0.0261 |
| GO:0035517 | PR-DUB complex | 2 | 0.0261 |
GO molecular function analysis revealed enrichment of key functional categories, included protein binding, cytoskeletal protein binding, and receptor binding. These terms showed fold-enrichment values ranging from approximately 4-7-fold and FDR values as low as 9.1 × 10-5, involving between 4 and 55 proteins per term (Table 5). These enriched functional categories highlight the dynamic interaction and signaling capabilities of MSC-EV cargo (Figure 7B).
| Term ID | Term description | Observed protein count | False discovery rate |
| GO:0005488 | Binding | 55 | 0.00044 |
| GO:0019899 | Enzyme binding | 20 | 0.0027 |
| GO:0005515 | Protein binding | 39 | 0.0041 |
| GO:0003682 | Chromatin binding | 9 | 0.0404 |
| GO:0019904 | Protein domain specific binding | 10 | 0.0404 |
| GO:1990782 | Protein tyrosine kinase binding | 5 | 0.0404 |
| GO:1990841 | Promoter-specific chromatin binding | 4 | 0.0404 |
GO biological process analysis revealed strong enrichment of pathways related to extracellular organization, wound healing, regulation of cell communication, and immune modulation. Several pathways showed high statistical significance (FDR < 0.001) and involved 3-27 proteins per term (Table 6), reflecting the diverse regenerative functions associated with MSC-EVs (Figure 7C).
| Term ID | Term description | Observed protein count | False discovery rate |
| GO:0001837 | Epithelial to mesenchymal transition | 12 | 5.06E-13 |
| GO:0048762 | Mesenchymal cell differentiation | 13 | 2.07E-11 |
| GO:0010717 | Regulation of epithelial to mesenchymal transition | 11 | 3.01E-11 |
| GO:0051893 | Regulation of focal adhesion assembly | 6 | 0.00002 |
| GO:1903690 | Negative regulation of wound healing, spreading of epidermal cells | 3 | 0.00016 |
| GO:1904261 | Positive regulation of basement membrane assembly involved in embryonic body morphogenesis | 3 | 0.00024 |
| GO:0001656 | Metanephros development | 6 | 7.17E-05 |
| GO:0045595 | Regulation of cell differentiation | 27 | 2.18E-11 |
| GO:0060677 | Ureteric bud elongation | 3 | 0.00034 |
| GO:0001822 | Kidney development | 10 | 1.04E-05 |
KEGG pathway analysis further revealed significant enrichment of biologically relevant pathways, including extracellular matrix-receptor interaction, phosphatidylinositol 3-kinase/protein kinase B (PI3K-AKT). signaling, focal adhesion, and cytokine-receptor interaction (FDR < 0.01). These pathways exhibited fold-enrichment values of approximately 3-8-fold and involved 7-8 proteins per pathway, highlighting the therapeutic potential of MSC-EVs in tissue repair and immunomodulation.
In this study, we analyzed MSC-EVs obtained from four bone marrow samples from healthy adult donors and identified a core set of 53 proteins, followed by comprehensive analysis of their molecular characteristics. Previous studies have reported that MSCs secrete multiple types of EVs that differ in size, marker expression, biochemical composition, and biological activity. Our findings are consistent with those reported in earlier studies[12,20-22].
EV cargo is not randomly distributed; rather, specific proteins and other biomolecules convey defined molecular signals to recipient cells. These nanoscale vesicles mediate intercellular communication by transferring proteins, mi
Several EV-associated proteins identified in our analysis are known to participate in organ development and tissue homeostasis, including processes related to kidney, ureter, and brain development. MSCs are widely recognized for their role in tissue repair, largely mediated through paracrine factors released via EVs. EVs contribute to tissue homeostasis and regeneration through their immunomodulatory properties and trophic factors, which are critical for the healing of injured tissues across multiple organ systems[25-28].
In our study, 15 of the 53 identified proteins were associated with regenerative functions, including angiogenesis, wound healing, and neural tissue repair. The protein cargo of EVs is highly cell type- and disease-specific, enabling EVs to deliver molecular signals that regulate biological processes in target tissues[29-31]. Advances in understanding MSC biology, together with improvements in EV isolation and characterization technologies, have significantly expanded the potential applications of EVs in regenerative medicine for restoring tissue function.
EVs contain a variety of cellular components, including proteins, nucleic acids (DNA and RNA), and lipids, allowing them to mediate therapeutic effects while avoiding some risks associated with cell-based therapies, such as uncontrolled cellular differentiation or mutations[32]. However, several important considerations remain. First, EVs carry and transfer genetic material, including DNA and RNA, which can influence the biological activity of recipient cells through intercellular communication and gene regulation. because EVs protect their genetic cargo within a lipid bilayer, they serve as stable and efficient vehicles for delivering genetic information to target cells[33].
Second, EV composition and functional properties may vary depending on donor-related factors, including age, health status, genetic background, and population diversity, as well as technical factors related to isolation methods[34]. These variables may influence the molecular characteristics and biological activity MSC-EVs. Finally, EV populations are inherently heterogene due to genetic, environmental, and lifestyle factors. This heterogeneity means that individual vesicles may differ in cargo composition and biological activity, which can affect the overall functional properties of EV preparations.
This study has several limitations. First, the sample size was small, as the analysis was based on EVs derived from only four donors. Second, given the genetic and ethnic diversity of the Indian population, the findings may not be fully generalizable. Third, EVs were isolated exclusively from BM-MSCs, and other potential MSC sources were not evaluated. Despite these limitations, this study provides valuable insights into the molecular characteristics of MSC-EVs.
In conclusion, proteomic analysis of MSC-EVs identified a conserved set of 53 proteins, including 15 proteins associated with regenerative functions. The relatively limited variation observed among EV profiles derived from BM-MSCs suggest the presence of a stable core EV proteome. These findings contribute to the growing understanding of MSC-EV biology and provide a foundation for future comparative studies involving larger and more diverse sample populations. Such studies will be important for advancing the development of EVs as emerging biomolecules with potential applications in both diagnostic and therapeutic settings.
| 1. | Bahari M, Mokhtari H, Yeganeh F. Stem Cell Therapy, the Market, the Opportunities and the Threat. Int J Mol Cell Med. 2023;12:310-319. [RCA] [PubMed] [DOI] [Full Text] [Cited by in RCA: 11] [Reference Citation Analysis (1)] |
| 2. | Kirkeby A, Main H, Carpenter M. Pluripotent stem-cell-derived therapies in clinical trial: A 2025 update. Cell Stem Cell. 2025;32:10-37. [RCA] [PubMed] [DOI] [Full Text] [Cited by in RCA: 118] [Reference Citation Analysis (0)] |
| 3. | Galipeau J, Sensébé L. Mesenchymal Stromal Cells: Clinical Challenges and Therapeutic Opportunities. Cell Stem Cell. 2018;22:824-833. [RCA] [PubMed] [DOI] [Full Text] [Full Text (PDF)] [Cited by in Crossref: 1510] [Cited by in RCA: 1360] [Article Influence: 170.0] [Reference Citation Analysis (5)] |
| 4. | Squillaro T, Peluso G, Galderisi U. Clinical Trials With Mesenchymal Stem Cells: An Update. Cell Transplant. 2016;25:829-848. [RCA] [PubMed] [DOI] [Full Text] [Cited by in Crossref: 1214] [Cited by in RCA: 1072] [Article Influence: 97.5] [Reference Citation Analysis (1)] |
| 5. | Trento C, Bernardo ME, Nagler A, Kuçi S, Bornhäuser M, Köhl U, Strunk D, Galleu A, Sanchez-Guijo F, Gaipa G, Introna M, Bukauskas A, Le Blanc K, Apperley J, Roelofs H, Van Campenhout A, Beguin Y, Kuball J, Lazzari L, Avanzini MA, Fibbe W, Chabannon C, Bonini C, Dazzi F. Manufacturing Mesenchymal Stromal Cells for the Treatment of Graft-versus-Host Disease: A Survey among Centers Affiliated with the European Society for Blood and Marrow Transplantation. Biol Blood Marrow Transplant. 2018;24:2365-2370. [RCA] [PubMed] [DOI] [Full Text] [Full Text (PDF)] [Cited by in Crossref: 71] [Cited by in RCA: 67] [Article Influence: 8.4] [Reference Citation Analysis (0)] |
| 6. | Trigo CM, Rodrigues JS, Camões SP, Solá S, Miranda JP. Mesenchymal stem cell secretome for regenerative medicine: Where do we stand? J Adv Res. 2025;70:103-124. [RCA] [PubMed] [DOI] [Full Text] [Full Text (PDF)] [Cited by in Crossref: 155] [Cited by in RCA: 110] [Article Influence: 110.0] [Reference Citation Analysis (2)] |
| 7. | Li P, Ou Q, Shi S, Shao C. Immunomodulatory properties of mesenchymal stem cells/dental stem cells and their therapeutic applications. Cell Mol Immunol. 2023;20:558-569. [RCA] [PubMed] [DOI] [Full Text] [Full Text (PDF)] [Cited by in Crossref: 207] [Cited by in RCA: 178] [Article Influence: 59.3] [Reference Citation Analysis (0)] |
| 8. | Szlasa W, Zendran I, Zalesińska A, Tarek M, Kulbacka J. Lipid composition of the cancer cell membrane. J Bioenerg Biomembr. 2020;52:321-342. [RCA] [PubMed] [DOI] [Full Text] [Full Text (PDF)] [Cited by in Crossref: 159] [Cited by in RCA: 304] [Article Influence: 50.7] [Reference Citation Analysis (0)] |
| 9. | Fan L, Wei A, Gao Z, Mu X. Current progress of mesenchymal stem cell membrane-camouflaged nanoparticles for targeted therapy. Biomed Pharmacother. 2023;161:114451. [RCA] [PubMed] [DOI] [Full Text] [Cited by in RCA: 33] [Reference Citation Analysis (3)] |
| 10. | Yokoi A, Ochiya T. Exosomes and extracellular vesicles: Rethinking the essential values in cancer biology. Semin Cancer Biol. 2021;74:79-91. [RCA] [PubMed] [DOI] [Full Text] [Cited by in Crossref: 19] [Cited by in RCA: 127] [Article Influence: 25.4] [Reference Citation Analysis (0)] |
| 11. | Di Bella MA. Overview and Update on Extracellular Vesicles: Considerations on Exosomes and Their Application in Modern Medicine. Biology (Basel). 2022;11:804. [RCA] [PubMed] [DOI] [Full Text] [Full Text (PDF)] [Cited by in Crossref: 2] [Cited by in RCA: 180] [Article Influence: 45.0] [Reference Citation Analysis (2)] |
| 12. | Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367:eaau6977. [RCA] [PubMed] [DOI] [Full Text] [Full Text (PDF)] [Cited by in Crossref: 9106] [Cited by in RCA: 8089] [Article Influence: 1348.2] [Reference Citation Analysis (16)] |
| 13. | Kandeel M, Morsy MA, Alkhodair KM, Alhojaily S. Mesenchymal Stem Cell-Derived Extracellular Vesicles: An Emerging Diagnostic and Therapeutic Biomolecules for Neurodegenerative Disabilities. Biomolecules. 2023;13:1250. [RCA] [PubMed] [DOI] [Full Text] [Cited by in RCA: 27] [Reference Citation Analysis (0)] |
| 14. | Clua-Ferré L, Suau R, Vañó-Segarra I, Ginés I, Serena C, Manyé J. Therapeutic potential of mesenchymal stem cell-derived extracellular vesicles: A focus on inflammatory bowel disease. Clin Transl Med. 2024;14:e70075. [RCA] [PubMed] [DOI] [Full Text] [Full Text (PDF)] [Cited by in Crossref: 57] [Cited by in RCA: 39] [Article Influence: 19.5] [Reference Citation Analysis (0)] |
| 15. | Liu TM. Mesenchymal stem cell-derived extracellular vesicles: Pioneering the next generation of biomedical applications. World J Stem Cells. 2025;17:108197. [RCA] [PubMed] [DOI] [Full Text] [Full Text (PDF)] [Cited by in RCA: 2] [Reference Citation Analysis (0)] |
| 16. | Ding Z, Greenberg ZF, Serafim MF, Ali S, Jamieson JC, Traktuev DO, March K, He M. Understanding molecular characteristics of extracellular vesicles derived from different types of mesenchymal stem cells for therapeutic translation. Extracell Vesicle. 2024;3:100034. [RCA] [PubMed] [DOI] [Full Text] [Full Text (PDF)] [Cited by in Crossref: 1] [Cited by in RCA: 40] [Article Influence: 20.0] [Reference Citation Analysis (0)] |
| 17. | Singh M, Tiwari PK, Kashyap V, Kumar S. Proteomics of Extracellular Vesicles: Recent Updates, Challenges and Limitations. Proteomes. 2025;13:12. [RCA] [PubMed] [DOI] [Full Text] [Cited by in RCA: 25] [Reference Citation Analysis (0)] |
| 18. | De Sousa KP, Rossi I, Abdullahi M, Ramirez MI, Stratton D, Inal JM. Isolation and characterization of extracellular vesicles and future directions in diagnosis and therapy. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2023;15:e1835. [RCA] [PubMed] [DOI] [Full Text] [Full Text (PDF)] [Cited by in Crossref: 61] [Cited by in RCA: 203] [Article Influence: 67.7] [Reference Citation Analysis (0)] |
| 19. | Li MY, Zhao C, Chen L, Yao FY, Zhong FM, Chen Y, Xu S, Jiang JY, Yang YL, Min QH, Lin J, Zhang HB, Liu J, Wang XZ, Huang B. Quantitative Proteomic Analysis of Plasma Exosomes to Identify the Candidate Biomarker of Imatinib Resistance in Chronic Myeloid Leukemia Patients. Front Oncol. 2021;11:779567. [RCA] [PubMed] [DOI] [Full Text] [Full Text (PDF)] [Cited by in Crossref: 5] [Cited by in RCA: 24] [Article Influence: 6.0] [Reference Citation Analysis (0)] |
| 20. | Lischnig A, Bergqvist M, Ochiya T, Lässer C. Corrigendum for "Quantitative Proteomics Identifies Proteins Enriched in Large and Small Extracellular Vesicles". Mol Cell Proteomics. 2023;22:100516. [RCA] [PubMed] [DOI] [Full Text] [Cited by in RCA: 3] [Reference Citation Analysis (0)] |
| 21. | Abyadeh M, Mirshahvaladi S, Kashani SA, Paulo JA, Amirkhani A, Mehryab F, Seydi H, Moradpour N, Jodeiryjabarzade S, Mirzaei M, Gupta V, Shekari F, Salekdeh GH. Proteomic profiling of mesenchymal stem cell-derived extracellular vesicles: Impact of isolation methods on protein cargo. J Extracell Biol. 2024;3:e159. [RCA] [PubMed] [DOI] [Full Text] [Full Text (PDF)] [Cited by in Crossref: 28] [Cited by in RCA: 25] [Article Influence: 12.5] [Reference Citation Analysis (0)] |
| 22. | Kowal J, Arras G, Colombo M, Jouve M, Morath JP, Primdal-Bengtson B, Dingli F, Loew D, Tkach M, Théry C. Proteomic comparison defines novel markers to characterize heterogeneous populations of extracellular vesicle subtypes. Proc Natl Acad Sci U S A. 2016;113:E968-E977. [RCA] [PubMed] [DOI] [Full Text] [Full Text (PDF)] [Cited by in Crossref: 3013] [Cited by in RCA: 2761] [Article Influence: 276.1] [Reference Citation Analysis (5)] |
| 23. | Andrade GM, Meirelles FV, Perecin F, da Silveira JC. Cellular and extracellular vesicular origins of miRNAs within the bovine ovarian follicle. Reprod Domest Anim. 2017;52:1036-1045. [RCA] [PubMed] [DOI] [Full Text] [Cited by in Crossref: 22] [Cited by in RCA: 40] [Article Influence: 4.4] [Reference Citation Analysis (1)] |
| 24. | Fazeli A, Godakumara K. The evolving roles of extracellular vesicles in embryo-maternal communication. Commun Biol. 2024;7:754. [RCA] [PubMed] [DOI] [Full Text] [Full Text (PDF)] [Cited by in RCA: 34] [Reference Citation Analysis (2)] |
| 25. | Varderidou-Minasian S, Lorenowicz MJ. Mesenchymal stromal/stem cell-derived extracellular vesicles in tissue repair: challenges and opportunities. Theranostics. 2020;10:5979-5997. [RCA] [PubMed] [DOI] [Full Text] [Full Text (PDF)] [Cited by in Crossref: 226] [Cited by in RCA: 221] [Article Influence: 36.8] [Reference Citation Analysis (5)] |
| 26. | Noiseux N, Gnecchi M, Lopez-Ilasaca M, Zhang L, Solomon SD, Deb A, Dzau VJ, Pratt RE. Mesenchymal stem cells overexpressing Akt dramatically repair infarcted myocardium and improve cardiac function despite infrequent cellular fusion or differentiation. Mol Ther. 2006;14:840-850. [RCA] [PubMed] [DOI] [Full Text] [Cited by in Crossref: 408] [Cited by in RCA: 359] [Article Influence: 18.0] [Reference Citation Analysis (1)] |
| 27. | de Windt TS, Vonk LA, Slaper-Cortenbach IC, van den Broek MP, Nizak R, van Rijen MH, de Weger RA, Dhert WJ, Saris DB. Allogeneic Mesenchymal Stem Cells Stimulate Cartilage Regeneration and Are Safe for Single-Stage Cartilage Repair in Humans upon Mixture with Recycled Autologous Chondrons. Stem Cells. 2017;35:256-264. [RCA] [PubMed] [DOI] [Full Text] [Cited by in Crossref: 200] [Cited by in RCA: 171] [Article Influence: 19.0] [Reference Citation Analysis (0)] |
| 28. | Bruno S, Grange C, Collino F, Deregibus MC, Cantaluppi V, Biancone L, Tetta C, Camussi G. Microvesicles derived from mesenchymal stem cells enhance survival in a lethal model of acute kidney injury. PLoS One. 2012;7:e33115. [RCA] [PubMed] [DOI] [Full Text] [Full Text (PDF)] [Cited by in Crossref: 519] [Cited by in RCA: 498] [Article Influence: 35.6] [Reference Citation Analysis (7)] |
| 29. | Pegtel DM, Peferoen L, Amor S. Extracellular vesicles as modulators of cell-to-cell communication in the healthy and diseased brain. Philos Trans R Soc Lond B Biol Sci. 2014;369:20130516. [RCA] [PubMed] [DOI] [Full Text] [Cited by in Crossref: 129] [Cited by in RCA: 184] [Article Influence: 16.7] [Reference Citation Analysis (0)] |
| 30. | Kalra H, Drummen GP, Mathivanan S. Focus on Extracellular Vesicles: Introducing the Next Small Big Thing. Int J Mol Sci. 2016;17:170. [RCA] [PubMed] [DOI] [Full Text] [Full Text (PDF)] [Cited by in Crossref: 463] [Cited by in RCA: 670] [Article Influence: 67.0] [Reference Citation Analysis (3)] |
| 31. | Tkach M, Théry C. Communication by Extracellular Vesicles: Where We Are and Where We Need to Go. Cell. 2016;164:1226-1232. [RCA] [PubMed] [DOI] [Full Text] [Cited by in Crossref: 3038] [Cited by in RCA: 2754] [Article Influence: 275.4] [Reference Citation Analysis (3)] |
| 32. | Phinney DG, Pittenger MF. Concise Review: MSC-Derived Exosomes for Cell-Free Therapy. Stem Cells. 2017;35:851-858. [RCA] [PubMed] [DOI] [Full Text] [Cited by in Crossref: 1518] [Cited by in RCA: 1384] [Article Influence: 153.8] [Reference Citation Analysis (5)] |
| 33. | Biller SJ, Ryan MG, Li J, Burger A, Eppley JM, Hackl T, DeLong EF. Distinct horizontal gene transfer potential of extracellular vesicles versus viral-like particles in marine habitats. Nat Commun. 2025;16:2126. [RCA] [PubMed] [DOI] [Full Text] [Full Text (PDF)] [Cited by in Crossref: 1] [Cited by in RCA: 17] [Article Influence: 17.0] [Reference Citation Analysis (0)] |
| 34. | Noren Hooten N, Byappanahalli AM, Vannoy M, Omoniyi V, Evans MK. Influences of age, race, and sex on extracellular vesicle characteristics. Theranostics. 2022;12:4459-4476. [RCA] [PubMed] [DOI] [Full Text] [Full Text (PDF)] [Cited by in Crossref: 3] [Cited by in RCA: 73] [Article Influence: 18.3] [Reference Citation Analysis (0)] |